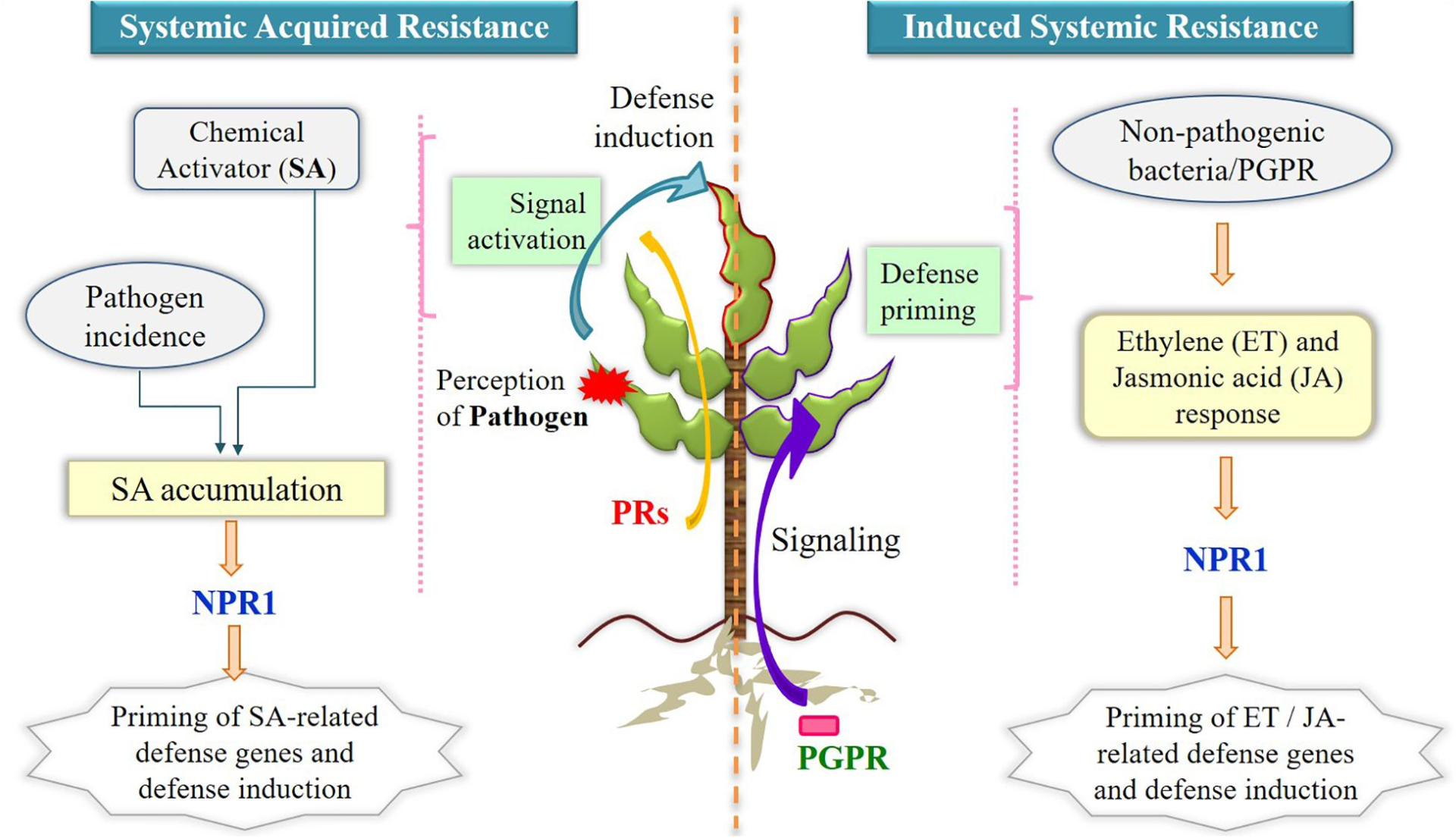
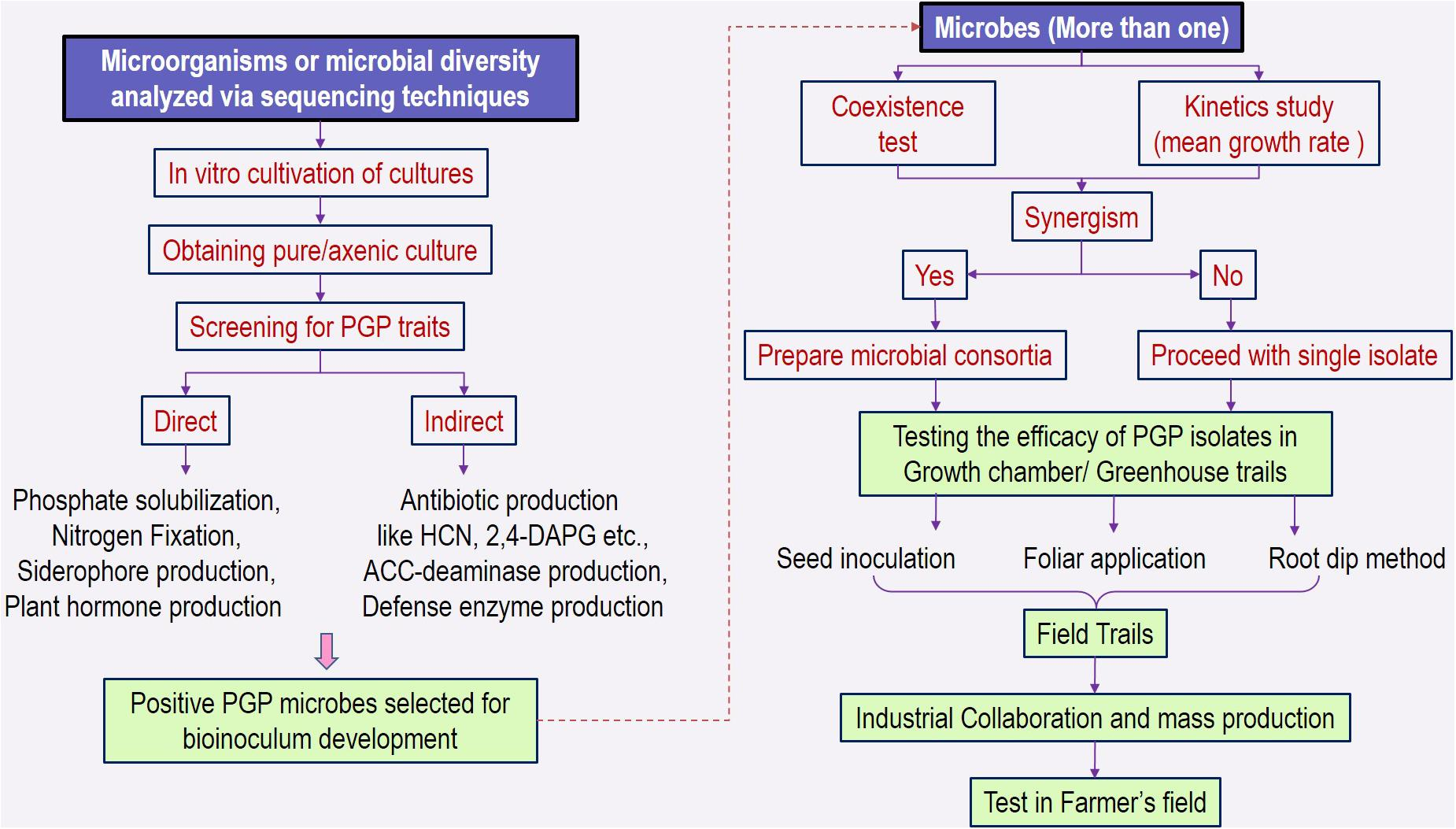
Revisiting Plant–Microbe Interactions and Microbial Consortia Application for Enhancing Sustainable Agriculture: A Review
Authors: Kanchan Vishwakarma, Nitin Kumar, Chitrakshi Shandilya, Swati Mohapatra, Sahil Bhayana and Ajit Varma
The present scenario of agricultural sector is dependent hugely on the use of chemical-based fertilizers and pesticides that impact the nutritional quality, health status, and productivity of the crops. Moreover, continuous release of these chemical inputs causes toxic compounds such as metals to accumulate in the soil and move to the plants with prolonged exposure, which ultimately impact the human health. Hence, it becomes necessary to bring out the alternatives to chemical pesticides/fertilizers for improvement of agricultural outputs. The rhizosphere of plant is an important niche with abundant microorganisms residing in it. They possess the properties of plant growth promotion, disease suppression, removal of toxic compounds, and assimilating nutrients to plants. Utilizing such beneficial microbes for crop productivity presents an efficient way to modulate the crop yield and productivity by maintaining healthy status and quality of the plants through bioformulations. To understand these microbial formulation compositions, it becomes essential to understand the processes going on in the rhizosphere as well as their concrete identification for better utilization of the microbial diversity such as plant growth–promoting bacteria and arbuscular mycorrhizal fungi. Hence, with this background, the present review article highlights the plant microbiome aboveground and belowground, importance of microbial inoculants in various plant species, and their subsequent interactive mechanisms for sustainable agriculture.

Figure 1. Associations in the rhizosphere between plant roots, microbes, and root exudates under biotic and abiotic influences.
Knowing the hugely diversified speciation, complexity in interactions, and structural composition of communities, the need of comprehending the root architectural biology and associated microbiome as an interactome becomes essential. The intertwining nature of host and microbes opens the possibility of numerous interactions such as plant root–root interactions and root–microbe interactions. Apart from this, root–nematode interactions also serve as an essential mode to understand the behavior of plants in response to such factors. Plant hosts and associated microbes possess inseparable ecological properties, which functions as metaorganism or holobiont (Hacquard and Schadt, 2015; Hacquard, 2016).
With the advancements in the techniques with respect to genome and proteome identification and analysis, studies are conducted to explore the mutual association between plant and microbes and understand related mechanisms for improved crop production (Bakker et al., 2013; Oldroyd, 2013). If the characteristics that are responsible for forming microbial community in the rhizosphere and its influence on plants are unraveled, these can be utilized for probable sustainable alternative in agroecosystem for enhanced stability and crop productivity in longer run (Quiza et al., 2015; Knapp et al., 2018). Hence, with this background, the review focuses on belowground microbial communities that start from their establishment to their interactions in the rhizosphere and mechanistic approaches and also highlights the aboveground plant microbiome.
A study conducted on microbiome of maize leaf across 300 plant cell lines showed that Sphingomonads and Methylobacteria are the predominant taxa (Wallace et al., 2018). It was also established that environmental factors play a major role in deriving microbial composition of the phyllosphere. Another study done by Steven et al. (2018) on apple flowers showed the dominance of Pseudomonas and Enterobacteriaceae taxa. Moreover, Pseudomonas has been observed to be an abundant genus in numerous studies conducted on flowers of apple, grapefruit, almonds, pumpkin, and tobacco (Aleklett et al., 2014). Recent studies were facilitated to assess the seed microbes, and it was observed that Firmicutes, Proteobacteria, Bacteroidetes, and Actinobacteria are the dominant ones (Liu et al., 2012; Barret et al., 2015; Rodríguez et al., 2018). The relation of seed microbiota has been seen with soil microbiota, and it is also evidenced that they can also be related to those of flowers and fruits (Compant et al., 2010; Glassner et al., 2018). The aboveground bacterial diversity originates from soil, seeds, and air followed by their inhabitation on or inside the plant tissues. Their existence on tissues is further shaped by various factors such as soil, environmental, and agricultural management practices. The strength of relationship between plant and its aboveground bacterial composition is specific to the host and the specific compartment where diversity exists; however, detailed knowledge of this relationship requires more research-based studies. These endophytes and aboveground microbiota are potentially known for promotion of plant growth, improvement of disease resistance, and alleviation of stresses (Hardoim et al., 2015; Vishwakarma et al., 2020).
Root exudation is defined as the secretion of several compounds of importance by the roots into the rhizosphere, for example, organic acids, sugars, amino acids, polyphenols, flavonoids, hormones, and nutrients, which act as source of nutrients for the microorganisms surrounding the roots (Mendes et al., 2013; Compant et al., 2019). This phenomenon is known as the rhizosphere effect. Nevertheless, the association of plant roots with microbiome involves the formation of selective niches for microbial development (Figure 2A). With the help of phytochemicals and root exudates, several microbial groups fail to grow in the rhizospheric niche. The population able to grow by utilizing root-secreted compounds forms a niche for themselves and also helps in recruiting other microbes by cross-feeding approach, thereby generating a new niche for rest of the microbes (Jacoby and Kopriva, 2019). The niche selection process is specific for the plant species and the compounds being secreted. For example, several secondary metabolites with defense properties such as benzoxazinoids discharged from the maize roots change the structure of root microbiome and influence the group of Actinobacteria and Proteobacteria the most (Hu et al., 2018). Moreover, the dynamics of structural composition of bacterial communities in the Avena barbata roots and their mechanisms were researched in a recent study (Zhalnina et al., 2018). It was observed that the amalgamation of root exudate composition and substrate selectivity significantly modified the assemblage of bacterial population in rhizosphere. Fitzpatrick et al. (2018) revealed various rhizobacterial species of Pseudoxanthomonas depicting differential patterns of occurrence across 30 angiospermic species. Moreover, the niche specifications and the huge diversity of the rhizospheric microbiota are also governed by the spatiotemporal organization of the rhizosphere and changes in physicochemical conditions (Vetterlein et al., 2020). On the whole, variety of plant species and related genotypes and components of root exudates affect the structure and alignment of rhizospheric microbiome (Vishwakarma et al., 2017a, b).

Figure 2. Interactions in the rhizosphere, (A) Plant–microbiome interactions: Plant roots secrete root exudates and phytochemicals that engage microbial populations in developing niches. Some metabolites filter out the unnecessary microbial strains occupied in niches (indicated by red cross), whereas some metabolites allow the different microbial population to coexist in same niche that may secrete compounds needed for growth of other microorganisms. (B) Root–root interactions: The neighboring plants may associate with other forming beneficial, as well as competing interactions by allelochemicals, root exudates, and volatile organic compounds. (C) Microbiome–plant interactions: Beneficial bacteria allow the promotion of plant growth by various mechanisms, such as making nutrients available by chelating them and transporting to plants (for example, siderophore-Fe transporter to carry utilizable iron); and producing phytohormones, such as indole acetic acid, secreted effectors, and antibiotics to protect plants from pathogens. AHL, N-acyl homoserine lactone; QSM, quorum-sensing molecules; VOCs, volatile organic compounds; Fe, iron; Cd, cadmium; Zn, zinc.
The internal colonization of roots also takes place by a variety of endophytic microbes. Their distribution in plants is dependent on several parameters such as the distribution of plant assets and the capability of endophytes in colonizing plants. One of the important and symbiotic root endophytes, Piriformospora indica, has been significantly used in agriculture for its function. The endophyte P. indica increases phosphorous (P) uptake and protects the crop from variety of stress factors (Lahrmann et al., 2013). It has been reported that a cyclophilin A–like protein from P. indica was overexpressed for protecting the tobacco plant against salt stress (Trivedi et al., 2013). It has been also observed that Azotobacter chrococcum can facilitate the modulation of P. indica physiology and helps in improving its nutrient acquisition through their synergistic action (Bhuyan et al., 2015).
Many endophytic fungi have been shown to exhibit chemotaxis for root-exuded chemicals. For instance, in a non-pathogenic Fusarium oxysporum when tested for activity against root knot nematode (Meloidogyne incognita) in tomato plants, it was found that the tomato exudates have facilitated the colonization of F. oxysporum, whereas it reduced the occurrence of nematode (Sikora and Dababat, 2007), suggesting that root exudates preferentially select the microbes in its vicinity. Nevertheless, root exudate–mediated chemotaxis also causes attraction for the pathogenic microbes. In a study by Gu et al. (2017), fine biochar was utilized to suppress bacterial wilt disease in tomato. The mechanism that biochar followed was absorption of root exudates that exerted strong chemotactic signal toward Ralstonia solanacearum, and as a result, its activity and swarming motility were suppressed. In a recent study, this bacterial pathogen has also been shown to follow chemotaxis for tomato root–exuded secondary metabolites (hydroxylated aromatic acids) (Hasegawa et al., 2019). Pratylenchus coffeae is an endoparasitic nematode that causes disruption of root tissues mechanically followed by invasion in plants (Das and Das, 1986). The molecular and gene expression studies on Pratylenchus coffeae have been conducted to specify the genes (related to cell wall degrading enzyme) regulated in the presence and absence of root exudates, and it was observed that their activity changed with respect to the host-specific root exudate components provided for the assay (Bell et al., 2019). The protozoan parasite Trypanosoma brucei generally displays its movement away from the other inhabited microbial groups; however, DeMarco et al. (2020) have recently observed their positive chemotactic effect toward the colonized area of Escherichia coli. It is due to the presence of attractant that is a soluble, diffusible signal dependent on actively growing E. coli.
Apart from this, different experiments were carried out to prove different evidence in relation to interactions between plant roots with differential niches. For instance, Semchenko et al. (2018) showed that vertically distributed roots are related to competitive interactions between plants rather than integral niche. Their results have shown that there is strong competition between the plant species, which spread their roots largely leading to the suppression of neighboring species, whereas species having deeper and less branched root system are extensively able to withstand such competition. Further, using genetically transformed plants, Weidlich et al. (2018) showed facilitative interactions between the roots of legume and non-legume species belowground. These interactions are limited not only to different species but also between the genotypes. Stepping from interactions between species to interactions between genotypes, Montazeaud et al. (2018) experimented on some species and observed the productivity of rice plants (Oryza sativa), which was grown in pairs, and it was observed that with increase in between-genotype distance, there was increase in mixture productivity in crops, which was attributed to resource-use complementarity. Moreover, mixing of two different species of trees was performed to explore soil by their fine roots. The species used were Acacia mangium and Eucalyptus grandis, where soil was more exploited by tree species as compared to the trees that were grown in the monoculture (Germon et al., 2018). These results further helped in supporting the importance of direct competition over the niche complementarity hypothesis.
It has been observed that the bacterial products/additives do not have clear understanding with respect to their adhesion, but adjustments in droplet size and rheological properties can be achieved by surfactants, which might help in improvement of adhesion to hydrophobic cuticular surfaces (Preininger et al., 2018). Improvement of adhesion of PGPRs to roots has been done by nanoparticles and humid environment provided by encapsulated PGPR macrobeads (Perez et al., 2018; Timmusk et al., 2018). Generally, yield of wheat in field studies is successfully increased by inoculation techniques adopted for inoculating seed, leaf, and soil of same PGPRs (Berger et al., 2018). Interference of seed inoculants with pesticides can be seen, but in such cases, seed inoculant colonizes the plants and activates microbial defense system, which include activation of plant immune response, biofilm production, etc. Development of new methods was done in addition to classical delivery approaches. Mitter et al. (2017) devised the concept of seed microbiome modulation. In this, flower spray inoculation was followed for achieving next-generation seeds colonized with endophytes and modulated seed microbiome. Colonization of germinated plants was done efficiently by inoculant strain, which displayed that the use of alternative approaches may lead to improvement of microbial inoculant performance under field conditions.
Microbial inocula, either single or consortia, have many advantages than limitations. These include their environment-friendly nature; they can restore soil fertility, improve/enhance nutrient availability, protect against biotic and abiotic stresses, increase soil microbial activity, decompose toxic substances, promote colonization of mycorrhizae and other useful microbes, help in recycling soil organic matter, increase plant defense and immunity for suppressing unwanted parasitic and pathogenic attacks, and carry out signal transduction and plant–microbe interactions. Each year, there is nearly 12% increase in demand for microbial inoculants because of the increasing cost of chemical fertilizers and demand for environment-friendly technologies in society (Calvo et al., 2014). PGPRs such as Azotobacter, Bacillus, Azospirillum, Pseudomonas, Burkholderia, Serratia, and Rhizobium species are now being commercially produced at a large scale (Parray et al., 2016), although different countries have their own rules for the use of these microbes based biofertilizers and biopesticides for agricultural practices (Bashan et al., 2014). The main obstacles are consistency, reliability, and shelf life of microbial inoculants under field conditions. Gram-positive bacteria have longer shelf-life in comparison to non–spore-forming gram-negative bacteria. However, studies have reported super-inoculants containing all the required characteristics of a microbial inoculant (Schoebitz et al., 2013). On the other hand, studies have also issued concern about some PGPRs that can be pathogenic to humans, for example, pathogenic Pseudomonas species and Burkholderia cepacia (Kumar et al., 2013). These species can be harmful to human, despite the PGP activity shown by them, and therefore before their commercial production, they should be addressed properly (Compant et al., 2010). More research is required before incorporating pathogenic PGPRs in sustainable agriculture. Many European and other countries such as the United States are reassessing the biosafety of PGPR-based biofertilizers. Studies have shown the effect of climate change on plant–microbe interactions; however, further studies are needed to know the full capabilities of PGPRs before their acceptance by government regulations, biofertilizer companies, and farmers. There can be the provision to make cost-effective technology of microbial consortium acceptance and utilization by the farmers in the future. There can be government-regulated outlets where biofertilizers/biopesticides with improved shelf life and stability should be provided to the farmers at subsidized rates with an opportunity to replace the old stored batch of inoculum with a fresh batch. The administrative bodies of agriculture-based towns can provide training to farmers highlighting the benefits, proper handling and usage, and their general guidelines. The schemes by the government can be launched to help farmers set up small production units in their area so as to regularize the inoculant production. It will certainly help them in overcoming shelf life, stability, and viable count problems by producing the inoculant as desired for the use.
Copyright © 2020 Vishwakarma, Kumar, Shandilya, Mohapatra, Bhayana and Varma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kanchan Vishwakarma, [email protected]; [email protected]; Ajit Varma, [email protected]
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Authors: Kanchan Vishwakarma, Nitin Kumar, Chitrakshi Shandilya, Swati Mohapatra, Sahil Bhayana and Ajit Varma
The present scenario of agricultural sector is dependent hugely on the use of chemical-based fertilizers and pesticides that impact the nutritional quality, health status, and productivity of the crops. Moreover, continuous release of these chemical inputs causes toxic compounds such as metals to accumulate in the soil and move to the plants with prolonged exposure, which ultimately impact the human health. Hence, it becomes necessary to bring out the alternatives to chemical pesticides/fertilizers for improvement of agricultural outputs. The rhizosphere of plant is an important niche with abundant microorganisms residing in it. They possess the properties of plant growth promotion, disease suppression, removal of toxic compounds, and assimilating nutrients to plants. Utilizing such beneficial microbes for crop productivity presents an efficient way to modulate the crop yield and productivity by maintaining healthy status and quality of the plants through bioformulations. To understand these microbial formulation compositions, it becomes essential to understand the processes going on in the rhizosphere as well as their concrete identification for better utilization of the microbial diversity such as plant growth–promoting bacteria and arbuscular mycorrhizal fungi. Hence, with this background, the present review article highlights the plant microbiome aboveground and belowground, importance of microbial inoculants in various plant species, and their subsequent interactive mechanisms for sustainable agriculture.
Introduction
Plants have dense inhabitation of the variety of microbes both belowground and aboveground that serve for their mutualistic benefits. The microbes that colonize the plants can be categorized into epiphytes that are present on the surface, endophytes that are located inside the plant tissues, phyllospheric that grow on leaf surfaces, and rhizospheric that inhabits into the soil close to the roots. Among them, rhizosphere is considered the most dynamic to significantly impact the nutritional status of plant and its growth (Bakker et al., 2013; Mendes et al., 2013; Lakshmanan et al., 2014). The term rhizosphere is defined as the narrow region of soil surrounding the roots and directly influenced by microbes and root secretions. The underground system comprises mainly soil and primary roots along with lateral developments and root hairs, which establish their interactions with countless microbial diversity in the rhizosphere, thereby significantly influencing the plant growth stages and resistance against variety of stresses (Figure 1) (Panke-Buisse et al., 2015; Bandyopadhyay et al., 2017). This whole system with plant roots interacting with the rhizomicrobiome constitutes the plant–root microbiome (Philippot et al., 2013).
Figure 1. Associations in the rhizosphere between plant roots, microbes, and root exudates under biotic and abiotic influences.
Knowing the hugely diversified speciation, complexity in interactions, and structural composition of communities, the need of comprehending the root architectural biology and associated microbiome as an interactome becomes essential. The intertwining nature of host and microbes opens the possibility of numerous interactions such as plant root–root interactions and root–microbe interactions. Apart from this, root–nematode interactions also serve as an essential mode to understand the behavior of plants in response to such factors. Plant hosts and associated microbes possess inseparable ecological properties, which functions as metaorganism or holobiont (Hacquard and Schadt, 2015; Hacquard, 2016).
With the advancements in the techniques with respect to genome and proteome identification and analysis, studies are conducted to explore the mutual association between plant and microbes and understand related mechanisms for improved crop production (Bakker et al., 2013; Oldroyd, 2013). If the characteristics that are responsible for forming microbial community in the rhizosphere and its influence on plants are unraveled, these can be utilized for probable sustainable alternative in agroecosystem for enhanced stability and crop productivity in longer run (Quiza et al., 2015; Knapp et al., 2018). Hence, with this background, the review focuses on belowground microbial communities that start from their establishment to their interactions in the rhizosphere and mechanistic approaches and also highlights the aboveground plant microbiome.
Aboveground Plant Microbiota
Unique environments for endophytic and epiphytic microbial diversities have been provided by different aboveground plant tissues such as vegetative foliar tissues, leaves, and floral parts, but the major differences in ecology of endospheric (endosphere is inside the environment of plant where microbes survive and may or may not be harmful to the plants; Hardoim et al., 2015; Compant et al., 2020) and phyllospheric (phyllosphere refers to the aerial region of the plant colonized by microbes) bacterial diversity exist. Systematic distribution of endophytes to different compartments such as stem, leaves, and fruits is facilitated by xylem (Compant et al., 2010), but it is observed that their entry to plant tissues can also take place through aerial parts such as fruits and flowers (Compant et al., 2011). Different compartments of plants possess distinct communities of endophytes, depending on source allocation of plant. The movement of phyllospheric bacteria is reportedly seen from soil environment that is driven by plant and various environmental parameters (Vorholt, 2012; Wallace et al., 2018). This leads to subsequent distribution of various microorganisms at genus and species level in endospheric and phyllospheric regions. For example, upon analyzing the structure of phyllosphere or carposphere of the grapevine, it was observed that Pseudomonas, Sphingomonas, Frigoribacterium, Curtobacterium, Bacillus, Enterobacter, Acinetobacter, Erwinia, Citrobacter, Pantoea, and Methylobacterium are predominant genera (Zarraonaindia et al., 2015; Kecskeméti et al., 2016), whereas when endophytes of grape berries were analyzed, the dominant genera found were Ralstonia, Burkholderia, Pseudomonas, Staphylococcus, Mesorhizobium, Propionibacterium, Dyella, and Bacillus (Campisano et al., 2014).A study conducted on microbiome of maize leaf across 300 plant cell lines showed that Sphingomonads and Methylobacteria are the predominant taxa (Wallace et al., 2018). It was also established that environmental factors play a major role in deriving microbial composition of the phyllosphere. Another study done by Steven et al. (2018) on apple flowers showed the dominance of Pseudomonas and Enterobacteriaceae taxa. Moreover, Pseudomonas has been observed to be an abundant genus in numerous studies conducted on flowers of apple, grapefruit, almonds, pumpkin, and tobacco (Aleklett et al., 2014). Recent studies were facilitated to assess the seed microbes, and it was observed that Firmicutes, Proteobacteria, Bacteroidetes, and Actinobacteria are the dominant ones (Liu et al., 2012; Barret et al., 2015; Rodríguez et al., 2018). The relation of seed microbiota has been seen with soil microbiota, and it is also evidenced that they can also be related to those of flowers and fruits (Compant et al., 2010; Glassner et al., 2018). The aboveground bacterial diversity originates from soil, seeds, and air followed by their inhabitation on or inside the plant tissues. Their existence on tissues is further shaped by various factors such as soil, environmental, and agricultural management practices. The strength of relationship between plant and its aboveground bacterial composition is specific to the host and the specific compartment where diversity exists; however, detailed knowledge of this relationship requires more research-based studies. These endophytes and aboveground microbiota are potentially known for promotion of plant growth, improvement of disease resistance, and alleviation of stresses (Hardoim et al., 2015; Vishwakarma et al., 2020).
Belowground Microbial Occurrence and Interactions
Microorganisms are ubiquitously present on the surfaces of plant along with their presence in the soil and are recruited by the plant from the surroundings, which then serve as microbial reservoirs (Hardoim et al., 2015). The root microbiome can be transferred in two different ways, i.e., horizontal and vertical. The dynamic communities of microbes associated with the plant roots generally undergo horizontal transfer, which means that they are enriched from the soil rich in diversified bacterial communities predominated by Acidobacteria, Bacteroidetes, Proteobacteria, Planctomycetes, and Actinobacteria (Fierer, 2017). The transfer of bacterial communities can also take place in vertical direction by seeds, representing an essential source of proliferating microbes from roots of a plant to its development (Hardoim et al., 2012). Distinct and interesting soil microbial niches are provided by the plant roots that allow their colonization in the rhizosphere and root, as well as aboveground areas to a certain limit (Hartmann et al., 2009). The narrow layer of soil in the vicinity of the plant roots (rhizosphere) is thought to be a highly active area for microbial movement, making it one of the most intricate environments (Hiltner, 1904). In a study, it was demonstrated by using culture-based technique, i.e., terminal restriction fragment length polymorphism, that abundant microbial community was present in the rhizosphere in comparison to the bulk soil in an extensive wheat cropping system (Donn et al., 2015).Root exudation is defined as the secretion of several compounds of importance by the roots into the rhizosphere, for example, organic acids, sugars, amino acids, polyphenols, flavonoids, hormones, and nutrients, which act as source of nutrients for the microorganisms surrounding the roots (Mendes et al., 2013; Compant et al., 2019). This phenomenon is known as the rhizosphere effect. Nevertheless, the association of plant roots with microbiome involves the formation of selective niches for microbial development (Figure 2A). With the help of phytochemicals and root exudates, several microbial groups fail to grow in the rhizospheric niche. The population able to grow by utilizing root-secreted compounds forms a niche for themselves and also helps in recruiting other microbes by cross-feeding approach, thereby generating a new niche for rest of the microbes (Jacoby and Kopriva, 2019). The niche selection process is specific for the plant species and the compounds being secreted. For example, several secondary metabolites with defense properties such as benzoxazinoids discharged from the maize roots change the structure of root microbiome and influence the group of Actinobacteria and Proteobacteria the most (Hu et al., 2018). Moreover, the dynamics of structural composition of bacterial communities in the Avena barbata roots and their mechanisms were researched in a recent study (Zhalnina et al., 2018). It was observed that the amalgamation of root exudate composition and substrate selectivity significantly modified the assemblage of bacterial population in rhizosphere. Fitzpatrick et al. (2018) revealed various rhizobacterial species of Pseudoxanthomonas depicting differential patterns of occurrence across 30 angiospermic species. Moreover, the niche specifications and the huge diversity of the rhizospheric microbiota are also governed by the spatiotemporal organization of the rhizosphere and changes in physicochemical conditions (Vetterlein et al., 2020). On the whole, variety of plant species and related genotypes and components of root exudates affect the structure and alignment of rhizospheric microbiome (Vishwakarma et al., 2017a, b).
Figure 2. Interactions in the rhizosphere, (A) Plant–microbiome interactions: Plant roots secrete root exudates and phytochemicals that engage microbial populations in developing niches. Some metabolites filter out the unnecessary microbial strains occupied in niches (indicated by red cross), whereas some metabolites allow the different microbial population to coexist in same niche that may secrete compounds needed for growth of other microorganisms. (B) Root–root interactions: The neighboring plants may associate with other forming beneficial, as well as competing interactions by allelochemicals, root exudates, and volatile organic compounds. (C) Microbiome–plant interactions: Beneficial bacteria allow the promotion of plant growth by various mechanisms, such as making nutrients available by chelating them and transporting to plants (for example, siderophore-Fe transporter to carry utilizable iron); and producing phytohormones, such as indole acetic acid, secreted effectors, and antibiotics to protect plants from pathogens. AHL, N-acyl homoserine lactone; QSM, quorum-sensing molecules; VOCs, volatile organic compounds; Fe, iron; Cd, cadmium; Zn, zinc.
The internal colonization of roots also takes place by a variety of endophytic microbes. Their distribution in plants is dependent on several parameters such as the distribution of plant assets and the capability of endophytes in colonizing plants. One of the important and symbiotic root endophytes, Piriformospora indica, has been significantly used in agriculture for its function. The endophyte P. indica increases phosphorous (P) uptake and protects the crop from variety of stress factors (Lahrmann et al., 2013). It has been reported that a cyclophilin A–like protein from P. indica was overexpressed for protecting the tobacco plant against salt stress (Trivedi et al., 2013). It has been also observed that Azotobacter chrococcum can facilitate the modulation of P. indica physiology and helps in improving its nutrient acquisition through their synergistic action (Bhuyan et al., 2015).
Many endophytic fungi have been shown to exhibit chemotaxis for root-exuded chemicals. For instance, in a non-pathogenic Fusarium oxysporum when tested for activity against root knot nematode (Meloidogyne incognita) in tomato plants, it was found that the tomato exudates have facilitated the colonization of F. oxysporum, whereas it reduced the occurrence of nematode (Sikora and Dababat, 2007), suggesting that root exudates preferentially select the microbes in its vicinity. Nevertheless, root exudate–mediated chemotaxis also causes attraction for the pathogenic microbes. In a study by Gu et al. (2017), fine biochar was utilized to suppress bacterial wilt disease in tomato. The mechanism that biochar followed was absorption of root exudates that exerted strong chemotactic signal toward Ralstonia solanacearum, and as a result, its activity and swarming motility were suppressed. In a recent study, this bacterial pathogen has also been shown to follow chemotaxis for tomato root–exuded secondary metabolites (hydroxylated aromatic acids) (Hasegawa et al., 2019). Pratylenchus coffeae is an endoparasitic nematode that causes disruption of root tissues mechanically followed by invasion in plants (Das and Das, 1986). The molecular and gene expression studies on Pratylenchus coffeae have been conducted to specify the genes (related to cell wall degrading enzyme) regulated in the presence and absence of root exudates, and it was observed that their activity changed with respect to the host-specific root exudate components provided for the assay (Bell et al., 2019). The protozoan parasite Trypanosoma brucei generally displays its movement away from the other inhabited microbial groups; however, DeMarco et al. (2020) have recently observed their positive chemotactic effect toward the colonized area of Escherichia coli. It is due to the presence of attractant that is a soluble, diffusible signal dependent on actively growing E. coli.
Root–Root Interactions
Because of the coexistence of different plants in the same soil, a competition is formed in the overlapping root systems for required resources that are limited in the soil. This coexistence has been thought relative to differentiation of niche because of different rooting patterns of plant species (Parrish and Bazzaz, 1976; Berendse, 1982). However, this theory supports competitive interactions occurring belowground. The surprising knowledge of coexistence also helps in showing the interactions that are competitive as well as facilitative between the co-occurring roots. The communication between roots of neighboring plants takes place by secretion of several signaling molecules such as root exudates and allelochemicals (Figure 2B) (Mommer et al., 2016a). Among them, allelopathy is the frequent communication process where phytotoxins such as catechin are released by plants. Catechin is capable of mediating both interspecific and intraspecific association by inhibiting growth of adjacent plant species, thereby enabling reduced competition and enhanced nutrient availability (Mommer et al., 2016b). Volatile organic compounds (VOCs) are also allelochemicals that mediate rhizospheric signaling by mycorrhiza networks among plants and increasing their transmission.Apart from this, different experiments were carried out to prove different evidence in relation to interactions between plant roots with differential niches. For instance, Semchenko et al. (2018) showed that vertically distributed roots are related to competitive interactions between plants rather than integral niche. Their results have shown that there is strong competition between the plant species, which spread their roots largely leading to the suppression of neighboring species, whereas species having deeper and less branched root system are extensively able to withstand such competition. Further, using genetically transformed plants, Weidlich et al. (2018) showed facilitative interactions between the roots of legume and non-legume species belowground. These interactions are limited not only to different species but also between the genotypes. Stepping from interactions between species to interactions between genotypes, Montazeaud et al. (2018) experimented on some species and observed the productivity of rice plants (Oryza sativa), which was grown in pairs, and it was observed that with increase in between-genotype distance, there was increase in mixture productivity in crops, which was attributed to resource-use complementarity. Moreover, mixing of two different species of trees was performed to explore soil by their fine roots. The species used were Acacia mangium and Eucalyptus grandis, where soil was more exploited by tree species as compared to the trees that were grown in the monoculture (Germon et al., 2018). These results further helped in supporting the importance of direct competition over the niche complementarity hypothesis.
Future Prospects, Challenges, and Limitations
To ensure long-term viability of microbial cells especially during storage and deliver sufficient viable number of cells to plants grown in fields, the development microbial formulations are needed. Unfortunate scene is that there is lack of suitable formulations for many microbes, in particular, the Gram-negatives (Berninger et al., 2018). Further limitation for viability in formulations is the toleration capacity of bacteria to low-humidity conditions (Köhl et al., 2011). Use of several compounds on the formulations might actually help in improvement of PGP effects. Experiments conducted for addition of LCOs isolated from rhizobia in the formulation or adaptation of growth medium of inoculants help in increasing exopolysaccharides and polyhydroxybutyrate content and increased PGP activities (Oliveira et al., 2017).It has been observed that the bacterial products/additives do not have clear understanding with respect to their adhesion, but adjustments in droplet size and rheological properties can be achieved by surfactants, which might help in improvement of adhesion to hydrophobic cuticular surfaces (Preininger et al., 2018). Improvement of adhesion of PGPRs to roots has been done by nanoparticles and humid environment provided by encapsulated PGPR macrobeads (Perez et al., 2018; Timmusk et al., 2018). Generally, yield of wheat in field studies is successfully increased by inoculation techniques adopted for inoculating seed, leaf, and soil of same PGPRs (Berger et al., 2018). Interference of seed inoculants with pesticides can be seen, but in such cases, seed inoculant colonizes the plants and activates microbial defense system, which include activation of plant immune response, biofilm production, etc. Development of new methods was done in addition to classical delivery approaches. Mitter et al. (2017) devised the concept of seed microbiome modulation. In this, flower spray inoculation was followed for achieving next-generation seeds colonized with endophytes and modulated seed microbiome. Colonization of germinated plants was done efficiently by inoculant strain, which displayed that the use of alternative approaches may lead to improvement of microbial inoculant performance under field conditions.
Microbial inocula, either single or consortia, have many advantages than limitations. These include their environment-friendly nature; they can restore soil fertility, improve/enhance nutrient availability, protect against biotic and abiotic stresses, increase soil microbial activity, decompose toxic substances, promote colonization of mycorrhizae and other useful microbes, help in recycling soil organic matter, increase plant defense and immunity for suppressing unwanted parasitic and pathogenic attacks, and carry out signal transduction and plant–microbe interactions. Each year, there is nearly 12% increase in demand for microbial inoculants because of the increasing cost of chemical fertilizers and demand for environment-friendly technologies in society (Calvo et al., 2014). PGPRs such as Azotobacter, Bacillus, Azospirillum, Pseudomonas, Burkholderia, Serratia, and Rhizobium species are now being commercially produced at a large scale (Parray et al., 2016), although different countries have their own rules for the use of these microbes based biofertilizers and biopesticides for agricultural practices (Bashan et al., 2014). The main obstacles are consistency, reliability, and shelf life of microbial inoculants under field conditions. Gram-positive bacteria have longer shelf-life in comparison to non–spore-forming gram-negative bacteria. However, studies have reported super-inoculants containing all the required characteristics of a microbial inoculant (Schoebitz et al., 2013). On the other hand, studies have also issued concern about some PGPRs that can be pathogenic to humans, for example, pathogenic Pseudomonas species and Burkholderia cepacia (Kumar et al., 2013). These species can be harmful to human, despite the PGP activity shown by them, and therefore before their commercial production, they should be addressed properly (Compant et al., 2010). More research is required before incorporating pathogenic PGPRs in sustainable agriculture. Many European and other countries such as the United States are reassessing the biosafety of PGPR-based biofertilizers. Studies have shown the effect of climate change on plant–microbe interactions; however, further studies are needed to know the full capabilities of PGPRs before their acceptance by government regulations, biofertilizer companies, and farmers. There can be the provision to make cost-effective technology of microbial consortium acceptance and utilization by the farmers in the future. There can be government-regulated outlets where biofertilizers/biopesticides with improved shelf life and stability should be provided to the farmers at subsidized rates with an opportunity to replace the old stored batch of inoculum with a fresh batch. The administrative bodies of agriculture-based towns can provide training to farmers highlighting the benefits, proper handling and usage, and their general guidelines. The schemes by the government can be launched to help farmers set up small production units in their area so as to regularize the inoculant production. It will certainly help them in overcoming shelf life, stability, and viable count problems by producing the inoculant as desired for the use.
Conclusion
With the increase in world population at alarming rate, there is a need to increase crop production to fulfill the global food requirements and at the same time enhance agricultural sustainability. Plant growth–promoting microbes, which are active constituents of biofertilizers and biopesticides, can be represented as a feasible alternative technology for enhancing plant yield and protecting against pathogens. The microbial inoculums possess the ability to positively impact the agriculture sector; however, plant selectivity along with organic and conventional management procedures also comes into play in shaping the rhizospheric microbiome structure, their concurrence, and subsequent effects. Since the microbial community structure in bulk and rhizosphere region frequently differs in their composition in various plant niches, it becomes necessary to reorganize the priorities of research toward isolating beneficial microbes and understanding the dynamics of their association with plants for enhanced crop productivity, quality, and agroecological sustainability. Despite some limitations of microbial consortia application, the measures to move past these limitations can be taken such as enhancement of shelf-life and viable load at the time of application, as well as developing faith in farmers for consistent utilization of inoculants in their fields. In the future, studies related to large-scale viable production of inoculant can be made using synergistic microbes proven to increase the crop productivity under conventional and organic agricultural practices.Author Contributions
AV and KV designed the structure of the manuscript. CS, NK, SM, and KV wrote the manuscript. CS, NK, KV, and SB prepared the tables, figures and arranged the references. KV and AV critically read and organized the manuscript. All the authors contributed to the article and approved the submitted version.Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.Acknowledgments
Authors would like to express thanks to ICAR-NASF, DST FIST, and DST Nano Mission for providing support.References
Click Download to read all the referencesCopyright © 2020 Vishwakarma, Kumar, Shandilya, Mohapatra, Bhayana and Varma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kanchan Vishwakarma, [email protected]; [email protected]; Ajit Varma, [email protected]
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.